


CYTOPROTECTIVE EFFECTS OF QUERCITIN AND SILYMARIN ON LIPO POLYSACCHARIDE – AND PARAQUAT-INDUCED CYTOTOXICITY IN ALVEOLAR MACROPHAGES: INFECTIOUS AND NON-INFECTIOUS CHALLENGES
HTML Full TextCYTOPROTECTIVE EFFECTS OF QUERCITIN AND SILYMARIN ON LIPOPOLYSACCHARIDE – AND PARAQUAT-INDUCED CYTOTOXICITY IN ALVEOLAR MACROPHAGES: INFECTIOUS AND NON-INFECTIOUS CHALLENGES
M. Khamisabadi 1, B. Lotfolahzadeh 1, A. Alenabi 1 and H. Malekinejad * 1, 2, 3
Department of Pharmacology and Toxicology 1, Faculty of Veterinary Medicine, P. O. Box: 1177, Urmia University, Urmia, Iran.
Food and Beverages Safety Research Center 2, Department of Pharmacology and Toxicology, Faculty of Pharmacy 3, Urmia University of Medical Sciences, Urmia, Iran.
ABSTRACT: Cytotoxicity of lipopolysaccharide (LPS) as a bacteria-related compound and paraquat (PQ) as a non-bacterial toxicant on freshly isolated alveolar macrophages (AMs) was studied. At the same time, preventive, protective and therapeutic effects of quercetin (QCN) against LPS-induced and silymarin (SMN) against PQ-produced cytotoxicity as known antioxidants with potent anti-inflammatory property were also investigated. Freshly isolated rat’s AMs were exposed to low, medium and high concentrations of LPS (1, 5 and 10 µg/ml), PQ (1, 5 and 10 µg/ml), QCN (1, 5 and 10 µg/ml) and SMN (25, 50 and 100 µg/ml), separately. After that, the effective concentration of LPS/PQ along with low, medium and high concentrations of QCN/SMN in the forms of preventive, protective and therapeutic was evaluated on cell viability, lactate dehydrogenase activity (LDH), nitric oxide content and total antioxidant capacity (TAC) of alveolar macrophages. Results showed that both LPS and PQ in a concentration-dependent manner reduced the cell viability and TAC values and elevated the NO content and LDH concentration. By contrast, QCN and SMN administration alone resulted in a non-significant increase of cell viability and significant (P<0.05) elevation of TAC values. Although, both QCN and SMN in all three proposed approaches attenuated the LPS/PQ-induced cytotoxicity respectively, a preventive form of the application, however, was found much appreciable than two other methods. Our data indicate that QCN and SMN could be novel and beneficial compounds to reduce the cytotoxic effects of LPS/PQ in AMs. Equally our findings suggest that AMs may be used as practical tools in further experimental research.
| Keywords: |
Alveolar macrophage, Cytotoxicity, Preventive, Protective, Therapeutic
INTRODUCTION: According to location, properties, and function two types of macrophages exist in the lungs. Alveolar macrophages (AM) and interstitial macrophages (IM) are located in the airway lumen and the parenchyma of the lungs, respectively.
AMs act as the first line host defense cells in the lungs via non-specific pathways including phagocytosis, the secretion of other chemicals such as nitric oxide (NO), tumor necrosis factor (TNF)-α and interferon (IFN-γ). Due to the facts that AMs activities are tightly regulated, hence they have been recognized as primary and pivotal pulmonary innate immune cells.
On the other hand, IMs function mainly with the release of certain cytokines such as interleukin, which are related to adaptive immune response 1. Flavonoids are phytochemicals that mainly are formed from the amino acids phenylalanine and tyrosine, and malonate. The nucleus of these compounds composes 3 basic rings (C6-C3-C6) and so-called flavan 2. Flavonoids also of having a defense role against insects in plants are playing a crucial role in the photosynthesis and phosphorylation of plants 3. Moreover, scavenging oxidants and in particular, absorbing the UV-generated reactive oxygen species (ROS) give important protective property for flavonoids in the various plants 4. There is plenty of evidence indicating different biological and medicinal activities of flavonoids including antioxidant, anti-thrombotic, anti-inflammatory, anti-atherogenic, anti-atherosclerotic, and cardioprotective effects 5.
One of the most abundant flavonoids is quercetin (QCN), which is found in the human diet and also in animals feed. During the last two decades, a broad range of its biological activities including its anti-inflammatory, antioxidant and immune-stimulatory effects have been frequently reported 6. Previous studies showed that QCN remarkably reduced the production of nitric oxide in endotoxin-stimulated microglia and protected from hydrogen peroxide-induced apoptosis in human neuronal SH-SY5Y cells 7.
Another well-known flavonoids, which during the last years has been used in human and animal medicine is silymarin (SMN), as the main component of S. marianum. Its hepatoprotective effect was the primary medicinal property, which let its worldwide use for the treatment of numerous liver disorders. SMN acts through the various pathways including antioxidant and anti-inflammatory activities, cell permeability regulator and membrane stabilizer, stimulating liver regeneration and inhibiting from cirrhosis.
Moreover, due to having the aforementioned properties, SMN currently is used as an effective and well-tolerated antidote against the hepatotoxic agents such as Amanita phalloide-produced toxic compounds 8. Other biological activities of SMN including its ability to inhibit the various cytochrome P450 isomers, its protective effects against Mycophenolate Mofetil-induced gastro-intestinal disorders and therapeutic effects on diabetes-produced consequences have been already studied 9, 10, 11. Despite the facts that several biological and medicinal properties of both flavonoids have been well documented, there is, however, little known about their effects on freshly isolated alveolar macrophages as a good experimental model of first-line respiratory defense against various agents. Hence, in the current study, we aimed to highlight possible cytoprotective effects of these two (QCN and SMN) flavonoids on lipopolysaccharide (LPS)- and paraquat (PQ)-induced cytotoxicity, respectively. The agents mentioned above were selected to play as good representatives for bacterial and non-bacterial related damages on AMs during respiratory disorders.
MATERIALS AND METHODS:
Chemicals: Silymarin (SMN, S 0292), quercetin (QCN, Q 0125), lipopolysaccharide (LPS, from Escherichia coli 0111: B4, L 2630), paraquat (PQ, 36541) were purchased from Sigma-Aldrich (Germany). Thiobarbituric acid, phosphoric acid (85%), trichloracetic acid (TCA), dimethyl sulfoxide (DMSO), ethyl acetate and ethanol were obtained from Merck (Germany). n-butanol was obtained from Carl Roth, GmbH Co. (Karlsruhe, Germany). Sulfanilamide was purchased from ACROS Organics, New Jersey (USA). Fetal bovine serum, Penicillin, Streptomycin, L-glutamine, and Dulbecco’s modified Eagle’s minimum were obtained from Gibco, Invitrogen, Carlsbad, CA, USA. MTT [(3-4, 5- dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide)] were purchased from Sigma Chemical Co., St Louis, MO, USA. All other chemicals were commercial products of analytical grade.
Isolation of Alveolar Macrophages: AMs were isolated from healthy and male Wistar rats (250 - 280 g). Animals were euthanized by overdose i.p. injection of Ketamin (80 mg/kg) and Xylazine (20 mg/kg). After exposing the trachea, it was cannulated, and immediately broncho-alveolar lavages were conducted using lavage solution (NaCl 140 mM, KCl 5 mM, phosphate buffer solution 2.5 mM, Hepes 10 mM, D-glucose 6 mM and ethylene diamine tetraacetic acid 0.2 mM). The used lavage solution kept at 37 °C during the entire period of cell collection. For each rat, 50 ml lavage solution in 5 times lavage processing was utilized. To minimize the heterogeneity of cells and to have a maximum population of AMs, the second fraction of broncho-alveolar lavage fluid was selected 12.
The collected lavage fluids were pooled and centrifuged at 500 ×g for 10 min, and the cell pellet was re-suspended in the cell culture medium of DMEM supplemented with 10% charcoal stripped Fetal Bovine Serum (FBS) and 1% Penicillin/ Streptomycin solution. The cells after counting with trypan blue method were allowed to adhere for 3h at 37 °C with 5% CO2. After 3 h incubation, non-adherent cells were aspirated, and complete fresh media was added to the attached cells.
Experimental Design: The isolated AMs were plated in 96-well plates at a density of 1.5 ×104 cells in 200 µl medium. The experiment was divided into two categories including screening the dose-dependency of all compounds and conducting experiments to show the beneficial effects of proposed flavonoids on PQ- and LPS-induced cytotoxicity as followings:
- Exposing the AMs against bacterial-related toxin (LPS, 1, 5 and 10 µg/ml);
- Exposing the AMs against PQ (1, 5 and 10 µg/ml) as a common and frequently occurring poisoning agent;
- Following a primary dose-dependent experiment for both LPS and PQ, in second step of study, the preventive, protective and therapeutic effects of QCN (1, 5 and 10 µg/ml) against LPS-induced and of SMN (25, 50 and 100 µg/ml) against PQ-induced cytotoxicity by using two known endpoints of colorimetric MTT and lactate dehydrogenase activity (LDH) measurement were investigated.
- To exclude any cytotoxic effects of both flavonoids on AMs, a concentration-dependent study using the above-mentioned concentrations of proposed flavonoids was also conducted. All three used concentration of tested chemicals were nominated as low (L), medium (M) and high (H) concentrations.
Our preliminary experiments revealed that for preventive studies, 6 h before the toxic agent exposure, proposed flavonoids could be added to the cultured AMs. Moreover, 6h after the exposure against toxic compounds was selected to study the therapeutic effects of flavonoids. Also of cytotoxicity assessments, to highlight any produced oxidative and nitrosative stress by used toxic agents and equally to clarify the antioxidant potency of used flavonoids, the total nitrite/nitrate concentration and total antioxidant capacity of treated and non-treated toxic agent-exposed AMs were determined.
Cytotoxicity Assays: AMs were grown on 96-well plates and challenged by adding increasing concentrations of LPS, QCN, PQ, SMN. For concentration-dependency and for 3 forms of preventive, protective and therapeutic studies, AMs exposed to various concentration of flavonoids for 24 h. Subsequently, the cytotoxic effect of LPS and PQ with and without flavonoids was evaluated by measuring lactate dehydrogenase (LDH) release and metabolic activity (MTT assay). The release of LDH was measured in the culture media using the CytoTox 96 non-radioactive cytotoxicity assay kit (Promega Corporation, WI, USA).
Furthermore, MTT reduction assay (Sigma-Aldrich, St Luis, Mo, USA) was performed to assess the metabolic activity of the challenged AMs. This assay measures the reduction of dimethyl thiazoldiphenyl tetrazolium bromide (MTT; stock solution 3 mg/mL dissolved in PBS) to formazan by the mitochondrial enzyme succinate dehydrogenase. Following exposing the cells against toxic compounds with or without flavonoids, the medium was discarded, and 0.2 ml MTT solution (3 mg/ml) was added to the cells. After 3h incubation at 37 ºC, the MTT solution was discarded, and the intracellular purple insoluble formazan was solubilized by adding 100 µL/well of lysis buffer (0.5% sodium dodecyl sulfate, 36 mMHCl and isopropanol acid). Following shaking on orbital shaker the optical density (OD) was measured at 570 nm using a microplate reader (BIO-RAD Model 3550). Cell viability was expressed as the percentage of non-treated controls as follows: A570 treated cells/A570 of control) × 100.
Nitric Oxide Measurement: The total nitrate/ nitrite content of AMs supernatant was measured according to the Griess reaction 13. In Griess reaction, nitric oxide rapidly converted into more stable nitrite, which in acidic environment nitrite is converted to HNO2. In reaction with sulphanilamide, HNO2 forms a diazonium salt, which reacts with N-(1-naphthyl) ethylenediamine. 2HCl to form an azo dye that can be detected by absorbing at 540 nm wavelength.
Assessment of Total Antioxidant Capacity (TAC): To determine the LPS and PQ impact and any beneficial effect of proposed flavonoids on total antioxidant capacity, the reducing capacity of AMs supernatant was measured. The assessment was performed based on ferric reduction anti-oxidant power (FRAP) assay 14. Briefly, at low pH which was provided using acetate buffer (300 mM, pH 3.6), reduction of FeIII-TPTZ (2, 4, 6-tri-2-pyridyl-1, 3, 5-triazin, Merck, Germany) complex to the ferrous form produces an intensive blue color that could be measured at 593 nm. The intensity of the complex following adding the appropriate volume of the serum to the reducible solution of FeIII-TPTZ is proportion to total reducing the power of the electron donating antioxidant. The aqueous solution of FeII(FeSO4.7H2O) was used as standard solutions.
Statistical Analyses: Mean and standard deviation of the measured parameters were calculated. The results of three independent experiments for each assessment were analyzed using Graph Pad Prism software (Version 2.01. Graph Pad Software Inc. San Diego, California, USA).
The comparisons between groups were made by analysis of variance (ANOVA) followed by Bonferroni post-hoc test. A P value <0.05 was considered significant.
RESULTS:
QCN and SMN Prevented from LPS and PQ-Reduced AMs Metabolic Activities: The metabolic activity of AMs was tested in the presence of QCN and LPS separately and in three distinct conditions by using the MTT test.
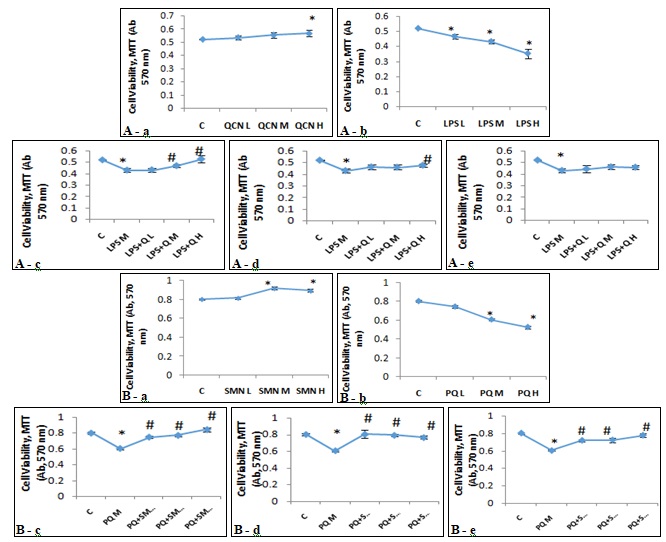
FIG. 1: EFFECTS OF (A): QUERCITIN IN THE PRESENCE AND ABSENCE OF LIPOPOLYSACCHARIDE AND (B): SILYMARIN IN THE PRESENCE OF PARAQUAT, ON VIABILITY OF ALVEOLAR MACROPHAGES (MTT ASSAY). The freshly isolated alveolar macrophages were exposed against various concentrations of QCN (A-a), LPS (A-b), SMN (B-a) and PQ (B-b) for 24 h in different approaches of preventive, protective and therapeutic models and cell viability was measured using MTT colorimetric assay. Asterisks are indicating significant differences between controls and all individual compounds and #s are presenting significant (P<0.05) differences between LPS/PQ-treated (Medium concentration) cells that received QCN/SMN at different concentrations and various models of preventive (A-c and B-c), protective (A-d and B-d) and/or therapeutic (A-e and B-e). L: low, M: Medium, H: High, QCN: Quercitin, LPS: Lipopolysaccharide, SMN: Silymarin and PQ: Paraquat.
Despite non-significant (P>0.05) elevation in MTT values with increasing concentration of QCN, LPS resulted in a marked and significant decline of cell viability at all three used concentrations. Cell viability was measured on cells, which exposed to the effective concentration of LPS in the presence of increasing concentrations of QCN. Only 6 h pre-incubation of AMs with QCN and after that exposing the cells against LPS resulted in concentration-dependent prevention of cell death. We found different reactions of the cells when the strategies of preventive, protection and treatment were applied. Both protective and therapeutic approaches only stopped the LPS-induced cell death, while the preventive policy even resulted in an increase of cell viability in a concentration-dependent manner Fig. 1A. We found the same reaction of AMs when exposed to SMN and PQ. SMN caused a significant increase in cell viability, and the highest rate of the cell population was found at a medium concentration of SMN (50 µg/ml). By contrast, PQ exposure resulted in a remarkable reduction in AMs metabolic activities. MTT assay revealed that SMN in all three approaches was able to combat the PQ-induced cytotoxicity Fig. 1B.
QCN and SMN Lowered the LPS-and PQ-Induced LDH Activities in AMs: AMs fate was also measured using LDH release assay, which is an indicator of cell membrane integrity.
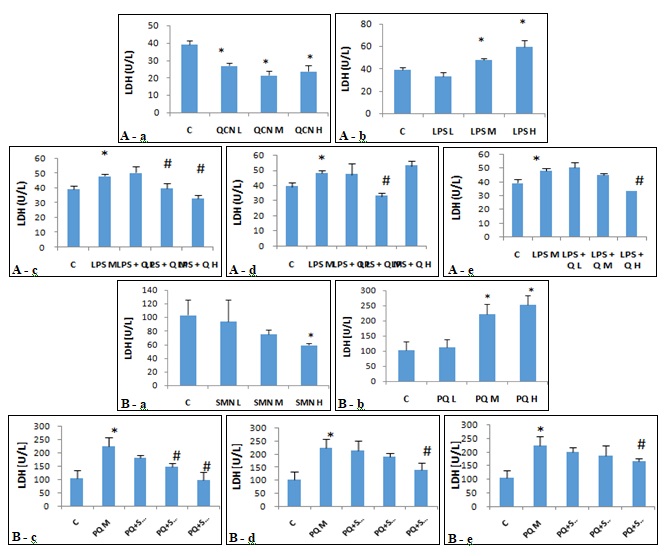
FIG. 2: EFFECTS OF (A): QUERCITIN IN THE PRESENCE AND ABSENCE OF LIPOPOLYSACCHARIDE AND (B): SILYMARIN IN THE PRESENCE AND ABSENCE OF PARAQUAT, ON RATE OF INJURED ALVEOLAR MACROPHAGES (LDH DETERMINATION). The freshly isolated alveolar macrophages were exposed against various concentrations of QCN (A-a), LPS (A-b), SMN (B-a) and PQ (B-b) for 24 h in different approaches of preventive, protective and therapeutic models and the rate of injured cells was measured by using the LDH assay. Asterisks are indicating significant differences between controls and all individual compounds and #s are presenting significant (P<0.05) differences between LPS/PQ-treated (Medium concentration) cells that received QCN/SMN at different concentrations and various models of preventive (A-c and B-c), protective (A-d and B-d) and/or therapeutic (A-e and B-e). L: low, M: Medium, H: High, QCN: Quercitin, LPS: Lipopolysaccharide, SMN: Silymarin and PQ: Paraquat.
LDH measurement showed that QCN in a non-concentration - dependent and LPS in a concentration-dependent fashion declined and elevated the LDH release from AMs into cell culture media, respectively. While both preventive and therapeutic approaches resulted in a considerable reduction of LDH release, application of protective policy showed only a significant (P<0.05) reduction of LDH release at a medium concentration of QCN Fig. 2A. LDH measurement in culture media from AMs, which exposed to SMN and PQ showed a significant and concentration-dependent reduction and elevation, respectively. Moreover, PQ-induced LDH release was declined by SMN in all three preventive, protective and therapeutic approaches. There were no significant (P>0.05) differences between tested approaches Fig. 2B.
QCN and SMN Reduced the NO Generation in AMs: The NO content of supernatant from AMs exposed to QCN and LPS individually was measured, and the obtained results showed that 24 h exposure of freshly isolated AMs to QCN resulted in non-significant changes in NO concentration, while LPS markedly at the same exposure time elevated NO level.

FIG. 3: EFFECTS OF (A): QUERCITIN IN THE PRESENCE AND ABSENCE OF LIPOPOLYSACCHARIDE AND (B): SILYMARIN IN THE PRESENCE AND ABSENCE OF PARAQUAT, ON THE GENERATION OF NITRIC OXIDE BY ALVEOLAR MACROPHAGES (GRIESS ASSAY). The freshly isolated alveolar macrophages were exposed against various concentrations of QCN (A-a), LPS (A-b), SMN (B-a) and PQ (B-b) for 24 h in different approaches of preventive, protective and therapeutic models and total nitrite/nitrate concentration was measured by Griess method. Asterisks are indicating significant differences between controls and all individual compounds and #s are presenting significant (P<0.05) differences between LPS/PQ-treated (Medium concentration) cells that received QCN/SMN at different concentrations and various models of preventive (A-c and B-c), protective (A-d and B-d) and/or therapeutic (A-e and B-e). L: low, M: Medium, H: High, QCN: Quercitin, LPS: Lipopolysaccharide, SMN: Silymarin and PQ: Paraquat.
While we failed to show any reduction of elevated NO concentration due to exposure to LPS in the supernatant of AMs, which were treated protectively and therapeutically with QCN, the LPS-induced NO generation was significantly (P<0.05) lowered when the cells were pretreated with QCN Fig. 3A.
To find out SMN and PQ concentration-dependent effect on the NO generation by isolated AMs, the NO content of exposed cells was measured and the obtained results indicate that, while PQ exposure resulted in marked elevation of NO concentration, SMN only at the highest tested concentration significantly (p<0.05) lowered the NO content in supernatant of AMs. Exposing AMs against the effective concentration of PQ and increasing concentrations of SMN in three various methods of preventive, protective and therapeutic showed that SMN at the highest given concentration and in the preventive approach resulted in a maximum reduction in PQ-induced NO concentration. Moreover, all three applied approaches lowered the PQ-increased NO concentration Fig. 3B.
Flavonoids Prevented from LPS- and PQ-Reduced TAC: The Total Antioxidant Capacity (TAC) of exposed cells to QCN and LPS was concentration-dependently elevated and reduced, respectively.
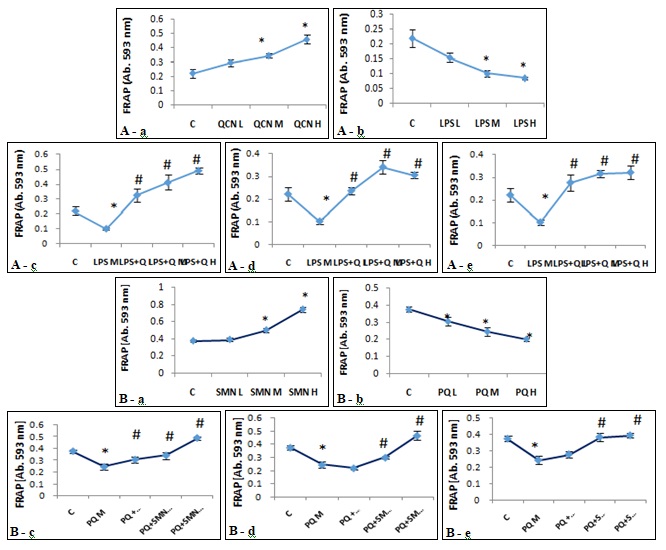
FIG. 4: EFFECTS OF (A): QUERCITIN IN THE PRESENCE AND ABSENCE OF LIPOPOLYSACCHARIDE AND (B): SILYMARIN IN THE PRESENCE AND ABSENCE OF PARAQUAT, ON TOTAL ANTIOXIDANT CAPACITY OF ALVEOLAR MACROPHAGES (FRAP ASSAY). The freshly isolated alveolar macrophages were exposed against various concentrations of QCN (A-a), LPS (A-b), SMN (B-a) and PQ (B-b) for 24 h in different approaches of preventive, protective and therapeutic models and total antioxidant capacity was measured by FRAP assay. Asterisks are indicating significant differences between controls and all individual compounds and #s are presenting significant (P<0.05) differences between LPS/PQ-treated (Medium concentration) cells that received QCN/SMN at different concentrations and various models of preventive (A-c and B-c), protective (A-d and B-d) and/or therapeutic (A-e and B-e). L: low, M: Medium, H: High, QCN: Quercitin, LPS: Lipopolysaccharide, SMN: Silymarin and PQ: Paraquat.
Pre-treatment of the freshly isolated AMs with QCN, not only prevented from LPS-reduced TAC values but also enhanced the antioxidant capacity significantly (P<0.05). Although the other two approaches also showed an increase of TAC, however the highest level of TAC was found in the preventive model of studies Fig. 4A.The freshly isolated AMs following 24 h incubation with SMN, revealed a concentration-dependent enhancement of TAC. At the same time, AMs also exposed to PQ, and the TAC values were determined in the supernatant. Results indicated that PQ exposure resulted in a significant (P<0.05) reduction of TAC values. Moreover, exposing the AMs against the effective concentration of PQ in the presence of increasing concentrations of SMN in three distinct approaches showed that all three forms not only recovered the PQ-reduced TAC values but also at the highest given concentration a significant elevation of TAC could be recorded when compared to the control cells Fig. 4B.
DISCUSSION: Flavonoids due to having phenolic hydroxyl moieties can scavenge the free radicals via electron donation 15. An increasing number of evidence indicates their beneficial effects on health, which mainly attribute to antioxidant and anti-inflammatory effects of these compounds. In the light of well-documented antioxidant and anti-inflammatory properties of flavonoids, we studied the possible role of two widely used flavonoids on two models of cytotoxicity, which may be counted as representative of bacterial- and non-bacterial-induced cytotoxicities in freshly isolated alveolar macrophages.
In this study, the AMs have been selected as an in-vitro model to study the importance of these cells as major effector cells against respiratory infections and also other toxicants, which may enter into the respiratory system. The reason for calling these cells as the first line of defense may be related to their multi-functional capabilities including phagocytosis, antigen presentation, production of different cytokines and enzymes 16. Another potency that makes AMs even more important is their migration capacity, which helps them to be accumulated at the most needed sites. Two well-known cytotoxicity tests have been used to study the concentration-dependent cytotoxicity of LPS and PQ on AMs. Both MTT assay by measuring the metabolic activity of cells and LDH test by assessing the rate of injured cells showed that LPS and PQ reduced the cell viability. To highlight the role of oxidative and nitrosative stress in the mechanism of cytotoxicity of LPS and PQ, produced the amount of nitric oxide and also total antioxidant capacity were measured.
Data showed that at least in part, the remarkable elevation of NO concentration and reduction of TAC play a crucial role in the cytotoxicity of both tested toxicants. NO despite being a messenger, effector and acting as a potent vaso-relaxant molecule in the body, the inflammation and sepsis is functioning as a deleterious molecule. Various cells including macrophages, smooth muscle cells, and hepatocytes in response to endogenous or exogenous exposure against stimulants such as bacterial toxins (for instance: LPS) inducible nitric oxide synthases are up-regulated and often followed by remarkable elevation of NO generation and ultimately major cellular injuries 17.
There is some other evidence supporting our findings that macrophages from rodent and human origin when exposed either to bacteria or bacteria-derived LPS, in a time and concentration-dependent manner enhanced the NO production 18, 19. Among the other molecular pathways, intracellular calcium elevation, activation of NOD2 and inhibition of LPS from binding to TLR4 have already been highlighted 20. PQ on the other hand although is not directly inhaled, it is however concentrated in the pulmonary system due to active polyamine uptake transport systems and causes acute lung injury. We and others recently showed that PQ poisoning in rats resulted in marked lipid peroxidation and also a significant increase of NO generation in the lungs 21, 22. We also clarified that part of the molecular pathway of PQ-induced respiratory toxicity is mediated in pulmonary macrophages and via involvement of PPARγ receptors 23. Therefore, the very first conclusion of the current study would be introducing AMs as a good model for investigations on respiratory inflammations and intoxications.
Our second finding indicates that both of tested flavonoids not only increased the antioxidant capacity but also slightly enhanced the cell viability as it was determined by MTT colorimetric and LDH assays. There is a various line of evidence indicating different ideas about the source of AMs including peripheral blood monocytes, the proliferation of AMs in the lung and proliferation in the in-vitro model in the presence of granulocyte-macrophage-colony-stimulating factor (GM-CSF) 24. Hence, our results may suggest that QCN and in particular SMN supplementation of growing medium might create a favorite condition for AMs to proliferate. Another aspect of the current study was to evaluate the most effective approach of administration of tested compounds. At first glance it becomes clear that both flavonoids in all three implemented approaches albeit with minor differences could be effective; however, we found that the most appreciable effects of every single flavonoid related to its preventive indication.
Since early time, there are plenty of interests to use the herbal medicines for various purposes, and recently much interest has been attracted on production and using of natural products both from research institutes and also from producers and users. Recent evidence indicates that there are epidemiological and experimental data showing that dietary polyphenols via antioxidant effects against low-density lipoprotein oxidation prevent atherosclerosis and also improve metabolic disorder and vascular inflammation 25. Flavonoids also in animal models showed anti-allergic effects through inhibition of chemical mediators including IL-4, Il-13 and IL-4-induced signal transduction 26, 27.
Concerning quercetin as the most abundant flavonoid in a wide range of fruits, vegetables and beverages due to showing anti-oxidant and anti-inflammatory effects, made the molecule a strong candidate for the prevention of cardiovascular diseases. Anti-inflammatory effects of quercetin are associated with its ability to reduce the production of TNF-α, IL-6, IL-8, IL-1β, interferon-γ-inducible protein-10, and cyclooxygenase(COX)-2 in human macrophages 28. At the same time, the preventive effect of SMN in the sepsis-induced lung injuries through regulation of inflammatory mediators and inhibition of neutrophils infiltration in human cases has been documented. There is recent evidence indicating that SMN protected from PQ-induced lung injuries through Nrf2-mediated pathway 29. Our findings in the current study consistently revealed that SMN remarkably reduced the PQ-induced NO production in all three approaches and by contrast increased significantly the PQ-reduced TAC values, suggesting that PQ-induced damages mainly mediated via oxidative and nitrosative stress, which was attenuated by SMN.
CONCLUSION: These results suggest that AMs might be a reliable and easily available model for testing the pathogenesis of respiratory toxicants and also for pulmonary inflammation inducers. Moreover, preventive effects of flavonoids and in particular two tested flavonoids in the various conditions of infection-related and toxicant-linked pulmonary inflammations were highlighted.
ACKNOWLEDGEMENT: Nil
CONFLICT OF INTEREST: The authors declare no conflict of interest.
REFERENCES:
- Byrne AJ, Mathie SA, Gregory LG and Lloyd CM: Pulmonary macrophages: key players in the innate defense of the airways. Thorax 2015; 70(12): 1189-1196.
- Pietta PG: Flavonoids as antioxidants. Journal of Natural Products 2000; 63(7): 1035-1042.
- Pietta PG and Simonetti P: Antioxidant food supplements. In: Packer L, Hiramatsu M and Yoshikawa T, editors: Antioxidant food supplements in human health. Eds.; Academic Press: San Diego 1999: 283-308.
- Salvamani S, Gunasekaran B, Shaharuddin NA, Ahmad SA and Shukor MY: Antiatherosclerotic effects of plant flavonoids. Biomed Research International 2014; (7): 480258. Doi:10.1155/2014/480258.
- Liao YR and Lin JY: Quercetin, but not its metabolite Quercetin-3-glucuronide, exerts prophylactic immuno-stimulatory activity and therapeutic anti-inflammatory effects on lipopolysaccharide-treated mouse peritoneal macrophages ex-vivo. Journal of Agriculture and Food Chemistry 2014; 62: 2872-2880.
- Suematsu N, Hosoda M and Fujimori K: Protective effects of quercetin against hydrogen peroxide-induced apoptosis in human neuronal SH-SY5Y cells. Neurosci Letters 2011; 504: 223-227.
- Bahmani M, Shirzad H, Rafieian S and Rafieian-Kopaie M: Silybum marianum: Beyond hepatoprotection. Journal of Evidence-Based Complementary and Alternative Medicine 2015; 20(4): 292-301.
- Malekinejad H, Rezabakhsh A, Rahmani F and Hobbenaghi R: Silymarin regulates the Cytochrome P450 3A2 and glutathione peroxides in the liver of strepto-zotocin-induced diabetic rats. Phytomedicine 2012; 19(7): 583-590.
- Malekinejad H, Sheikhzadeh S and Hobbenaghi R: Silymarin attenuated the Mycophenolate Mofetil-induced gastrointestinal disorders in rats. Avicenna Journal of Phytomedicine 2014; 4: 170-181.
- Malekinejad H, Rokhsartalab-Azar S, Hassani-Dizaj S, Alizadeh-Fanalous H, Rezabakhsh A and Amniattalab A: Effects of silymarin on the pharmacokinetics of atorvastatin in diabetic rats. European Journal of Drug Metabolism and Pharmacokinetics 2014; 39(4): 311-320.
- Laplante C and Lemaire I: Interactions between alveolar macrophage subpopulations modulate their migratory function. The American Journal of Pathology 1990; 136: 199-206.
- Green LC, Wagne AD, Glogowski J, Skipper PL, Wishnok JS and Tannenbaum SR: Analysis of nitrate, nitrite and [15N] nitrate in biological fluids. Analytical Biochemistry 1982; 126(1): 131-138.
- Benzie IF and Strain JJ: Ferric reducing/antioxidant power assay: direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods in Enzymology 1999; 299:15-27.
- Furusawa M, Tanaka T and Ito T: Antioxidant activities of hydroxyl flavonoids. Journal of Health Science 2005; 51: 376-378.
- Mutua MP, Muita SM and Gicheru M: Protective roles of free avian respiratory macrophages in captive birds. Biological Research 2016; 49: 29. doi: 10.1186/s40659-016-0090-7.
- Shen SC, Lee WR, Lin HY, Huang HC, Ko CH, Yang LL and Chen YC: In-vitro and in-vivo inhibitory activities of rutin, wogonin and quercetin on lipopolysaccharide-induced nitric oxide and prostaglandin E2 production. European Journal of Pharmacology 2002; 446(1-3): 187-194.
- Landes MB, Rajaram MVS, Nguyen H and Schlesinger LS: Role for NOD2 in Mycobacterium tuberculosis-induced iNOS expression and NO production in human macrophages. Journal of Leukocyte Biology 2015; 97(6): 1111-1119.
- Zhang X, Feng J, Zhu P and Zhao Z: Ketamine inhibits calcium elevation and hydroxyl radical and nitric oxide production in lipopolysaccharide-stimulated NR8383 alveolar macrophages. Inflammation 2013; 36(5): 1094-1100.
- Joh EH, Gu W and Kim DH: Echinocystic acid ameliorates lung inflammation in mice and alveolar macrophages by inhibiting the binding of LPS to TLR4 in NF-kB and MAPK pathways. Biochemical Pharmacology 2012; 84: 331-340.
- Malekinejad H, Rezabakhsh A, Rahmani F and Razi M: Paraquat exposure up-regulates Cyclooxygenase-2 in the lungs, liver and kidneys in rats. Iranian Journal of Pharmaceutical Research 2013; 12(4): 887-896.
- Aires RD, Capettini LSA, Silva JF, Rodriques-MachadoMda G, Pinho V, Teixeira MM, Cortes SF and Lemos VS: Paraquat poisoning induces TNF-α-dependent iNOS/NO mediated hyporesponsiveness of the aorta to vasoconstrictors in rats. PLoS One 2013; 8(9): e73562. doi:10.1371/journal.pone.0073562.
- Malekinejad H, Khoramjouy M, Hobbenaghi R and Amniattalab A: Atorvastatin attenuates the paraquat-induced pulmonary inflammation via PPARγ receptors: A new indication for atorvastatin. Pesticide Biochemistry and Physiology 2014; 114: 79-89.
- Nakata BK, Gotoh H, Watanabe J, Uetake T, Komuro I, Yuasa K, Watanabe S, Ieki R, Sakamaki H, Akiyama H, Kudoh S, Naitoh M, Satoh H and Shimada K: Augmented proliferation of human alveolar macrophages after allogeneic bone marrow transplantation. Blood 1999; 93(2): 667-673.
- Kishimoto Y, Tani M and Kondo K: Pleiotropic preventive effects of dietary polyphenols in cardiovascular diseases. European Journal of Clinical Nutrition. 2013; 67: 532-535.
- Das M, Ram A and Ghosh B: Luteolin alleviates bronchoconstriction and hyper airway reactivity in ovalbumin-sensitized mice. Inflammation Research 2003; 52: 101-106.
- Leemans J, Cambier C, Chandler T, Billen F, Clercx C and Kirschvink N: Prophylactic effects of omega-3 polyunsaturated fatty acids and luteolin on airway hyper responsiveness and inflammation in cats with experimentally-induced asthma. The Veterinary Journal 2010; 184(1): 111-114.
- Overman A, Chuang CC and McIntosh M: Quercetin attenuates inflammation in human macrophages and adipocytes exposed to macrophage-conditioned media. International J. of Obesity (Lond) 2011; 35(9): 1165-1172.
- Toklu HZ, Akbay TT, Velioglu-Ogunc A, Erkan F, Gedik N and Keyer-Uysal M: Silymarin, the antioxidant component of Silybum marianum, prevents sepsis-induced acute lung and brain injury. Journal of Surgery Research 2008; 145(2): 214-222.
- Zhao F, Shi D, Li T, Li L and Zhao M: Silymarin attenuates paraquat-induced lung injury via Nrf2-mediated pathway in-vivo and in-vitro. Clinical and Experimental Pharmacology and Physiology 2015; 42: 988-998.
How to cite this article:
Khamisabadi M, Lotfolahzadeh B, Alenabi A and Malekinejad H: Cytoprotective effects of quercitin and silymarin on lipopolysaccharide- and paraquat-induced cytotoxicity in alveolar macrophages: infectious and non-infectious challenges. Int J Pharmacognosy 2018; 5(9): 571-80. doi link: http://dx.doi.org/10.13040/IJPSR.0975-8232.IJP.5(9).571-80.
This Journal licensed under a Creative Commons Attribution-Non-commercial-Share Alike 3.0 Unported License.
Article Information
4
571-580
835
1425
English
IJP
M. Khamisabadi, B. Lotfolahzadeh, A. Alenabi and H. Malekinejad *
Food and Beverages Safety Research Center, Urmia University of Medical Sciences, Urmia, Iran.
hassanmalekinejad@yahoo.com
05 June 2018
03 July 2018
09 July 2018
10.13040/IJPSR.0975-8232.IJP.5(9).571-80
01 September 2018