ASSESSMENT OF MALATHION INDUCED TOXICITY IN DATTAPHRYNUS MELANOSTICTUS TADPOLES: A BIOCHEMICAL INVESTIGATION
HTML Full TextASSESSMENT OF MALATHION INDUCED TOXICITY IN DATTAPHRYNUS MELANOSTICTUS TADPOLES: A BIOCHEMICAL INVESTIGATION
R. M. Kartheek 1, M. David * 2, G. P. Manjunath 2, P. Lokeshkumar 2 and D. Mahantesh 2
Department of Zoology 1, JSS Banashankari Arts Commerce and SK Gubbi Science College, Dharwad - 580004, Karnataka, India.
Environmental Toxicology and Molecular Biology Laboratory 2, Department of PG Studies and Research in Zoology, Karnatak University, Dharwad - 580003, Karnataka, India.
ABSTRACT: Environmental pollutants apart from other factors like habitat loss and interventions of invasive species, is regarded as a likely cause towards a worldwide decline in amphibian population. The present study attempts to elucidate the toxicity of commercially formulated malathion (MAT) by investigating biochemical aspects in Dattaphrynus melanostictus tadpoles. Three sublethal concentrations of MAT (1.0, 1.8, and 2.5 mg/l) were considered to which the tadpoles (Gosner stage 27) were exposed for five days. The outcome of the present investigation revealed a significant decline (P<0.05) in activities of catalase, superoxide dismutase, glutathione peroxidase while suggesting a significant elevation in lipid peroxidation. The inhibition acetylcholinesterase activity confirmed MAT as anticholinesterase product. The overall outcome of the present investigation suggests the toxic potentials of MAT; which could have possibly resulted in compromised antioxidant status and neurobiochemical makeup of the exposed tadpoles. Thus, based on the results obtained, it could be ascertained that the commercial grade MAT may pose a potential threat to the tadpoles of D. melanostictus under the selected sublethal concentrations. The study further validates the feasibility to measure the intensity of aquatic pollution in the course of regulatory surveillance and monitoring the waters with suspected organophosphate contamination.
| Keywords: |
Acetylcholinesterase, Oxidative stress, Pesticide toxicity, Pollution and tadpoles
INTRODUCTION: Large scale anthropogenic activities have been associated with the drastic decline of amphibian populations globally 1, 2, 3. Evaluation by the International Union for Conservation of Nature, category for vulnerable or critically endangered species suggested that 32.5% of total amphibian species have declined in terms of their number, which is far critical than for birds and mammals 4, 5.
Amphibians, unlike other animals, constitute a unique group among many ecosystems due to their active and multiple roles as, prey, predators and herbivores 6, 7. Their contribution to trophic dynamics makes them one of the crucial features in determining the survival ability of other organisms through food chain 8, 9. Hence, their existence at a certain population ratio could be considered as accountable for the continuity of other species as well. Even though the loss of habitat is considered to be the primary reason behind amphibian decline 10, the role of pesticide contamination in freshwater habitats often questions its contribution in survival rate and reproduction of anurans 11.
In addition to this, tadpoles are known to complete a part of their life cycle (larval development) in an aquatic medium like ponds and lakes to which the pesticidal effluents from agricultural runoffs often find their way 12, 13, 14. The rich permeability of skin and egg, which often results in getting absorbed, persist, and bioaccumulate further explains the vulnerability of tadpoles to environmental xenobiotics 15.
A number of factors like morphological deformities, compromised reproducing ability, immune-suppression, and reduction in growth and development has indicated the potential risk of pesticide contamination against anurans 16. The use of integrated biomarker approaches for studying the inter-cascading changes in biochemistry has become an advanced strategy for reporting the overall health of tadpoles under the toxicological point of view 17, 18. Amphibian susceptibility to insecticides has been very well acknowledged in the past 15 and proven to be critical enough for compromising growth and development 19.
These insecticides are further known to indirectly increase susceptibility to parasite infection by decreasing the activity patterns in tadpoles 20. Since tadpoles are capable of avoiding free-swimming parasites by either moving away or swimming in erratic patterns 21, the response through means of locomotory perception upon suffering pesticide-induced physiological trauma is highly critical in determining their survival tendencies 22 and hence, cannot be overlooked.
Previous reports have suggested the involvement of toxicants in imparting oxidative stress by random generation of reactive oxygen species 23, 24, 25 and also known to pose catastrophic potentials against neurotransmission in tadpoles that may result in impaired swimming abilities 26. However, literature support on evaluations of MAT toxicity in tadpoles of Dattaphrynus melanostictus is found to be limited. Even though D. Melanostictus is found being fairly resistant as compared to other species of tadpoles 27, susceptibility in terms of antioxidant enzyme status and neurobiochemical activity cannot be ignored under the declining trend of its population. In addition to this, the effect of MAT on activities of free oxyradical scavenging enzymes and neurotransmission capabilities lacks in literature. Therefore, an attempt has been made in the present study to investigate the toxic potentials of sublethal concentrations of commercially formulated MAT on D. melanostictus tadpoles.
MATERIALS AND METHODS:
Toxicant Selected and Test Solutions: Commercial grade malathion of 50% EC (MAT) was selected as the toxicant for the present study and was procured from the local market (Dharwad, Karnataka, India). The stock solution was prepared by dissolving 1 gram of MAT in 100 ml of double distilled water. The requisite test concentrations were freshly prepared by diluting the stock solution before the initiation of toxicity studies.
Procurement and Maintenance of Tadpoles: Five hundred and fifty tadpoles of Dattaphrynus melanostictus (Stage 20) were collected from uncontaminated ponds located in Karnatak University campus, Dharwad city (Karnataka, India) and were transported to the laboratory with care. The tadpoles were acclimatized to laboratory conditions, during which they were fed with plant origin feed and boiled spinach ad libitum. The water in aquaria was renewed every two days, and excess of food and feces were removed. For exposure studies, tadpoles were divided into four groups, namely, control (C) and Exposure 1 (E1), Exposure 2 (E2), Exposure 3 (E3). The groups namely C, E1, E2, and E3 received MAT concentrations of 0.0, 1.0, 1.8 and 2.5 mg/l respectively. Each group consisted of 10 tadpoles (n=10) and were maintained in triplicates. Tadpoles were transferred to glass aquaria only after thorough inspection and identification of its stage (Stage 27) according to Gosner 28. Each glass aquaria consisted of 10 liters of water, which was dechlorinated through aeration. All the experiments were carried out at 25 ± 2 ºC with a light-dark cycle of 14:10 hours, which was being maintained throughout the completion of the investigation. Besides, temperature, pH, and dissolved oxygen levels were monitored daily Table 1. The mean values of individual group were taken into account for the present study.
Antioxidant Assay: The antioxidant status was determined by analyzing variation in the activity of antioxidant enzymes. The antioxidant enzymes, catalase, superoxide dismutase, glutathione peroxidase, and lipid peroxidation level was determined by the methods of Luck 29, Kakkar et al.,30, Paglia and Valentine 31 and Buege and Aust 32 respectively.
TABLE 1: SHOWING VALUES FOR QUALITY ASSESSMENT OF WATER USED IN THE PRESENT INVESTIGATION
| Parameter | Values obtained |
| Temperature | 24 ± 1 ºC |
| pH | 7.1 ± 0.3 |
| Dissolved oxygen | 6.1 ± 0.4 mg/L |
| Total Hardness | 37.3 ± 3.1 mg as CaCO3/L |
| Salinity | Nil |
| Specific gravity | 1.003 |
| Calcium | 21.31± 0.27 mg/L |
| Phosphate | 0.9 ± 0.04 mg/L |
| Magnesium | 0.85 ± 0.3 mg/L |
Determination of AChE Activity: The activity of acetylcholinesterase was determined by the methodology as described by Ellman et al. 33
Statistical Analysis: The enzymatic activities of neurobiochemical and antioxidant status are reported as the mean ± standard error of the mean (SEM) obtained from triplicates. The data were subjected to one-way analysis of variance and further subjected to Tukey’s test for post hoc analysis by defining the significance level at Ρ<0.005.
Ethical Committee: All procedures implemented in the present study were in accordance with the guidelines of the Institutional Animal Ethics Committee (IAEC). The animals subjected to experimentation were handled as per the guidelines issued by the Committee for the Purpose of Control and Supervision of Experiments for Animals (CPCSEA), New Delhi, India.
RESULTS: The results from the present investigation suggested the toxic insult due to MAT intoxication which was by inducing oxidative stress due to variations in all the considered antioxidant enzyme activities and curtailing acetylcholi-nesterase activity in all groups affecting the biochemistry exposed tadpoles. The variation of catalase activity in‘E1’ was noticed which showed a variation of +18.43%, followed by the change of -28.49% and -60.41% in ‘E2’ and ‘E3’ respectively when compared with control ‘C’ Fig. 1.
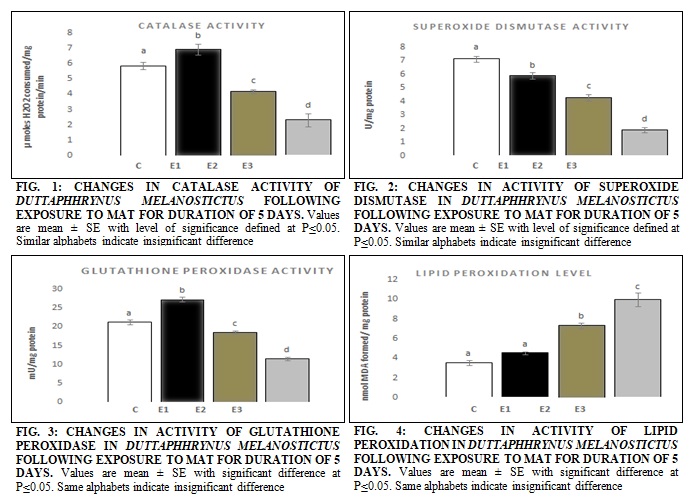
The decline in the activity of SOD was observed in a uniform trend which demonstrated the continuous destruction of its enzymatic activity. The percent decline in SOD activity was -17.13%, -39.60% and -73.73% in‘E1’, ‘E2’ and ‘E3’ respectively as compared to ‘C’ Fig. 2. A significant difference was also noticed in the activity of GPx in the tadpoles of different groups, Fig. 3. The percent change in expression patterns of GPx was found to be; +28.27%, -12.48% and -45.85% in ‘E1’, ‘E2’ and ‘E3’ respectively when compared with ‘C.’
The lipid peroxidation was found to elevate under the influence of MAT, and the variations of the same have been presented in Fig. 4. The increase in lipid peroxidation of exposed tadpoles witnessed in the present study was continuous, and the increase in MDA formation has been presented in Fig. 4. The significant difference in ‘E2’ and ‘E3’ was noticed in exposed tadpoles unlike ‘E1’. Upon correlation, the results indicated the frequency of damage to be highest in tadpoles belonging to ‘E3’ which endured the highest amount of toxicity caused by the maximum concentration of MAT after 5 days. AChE activity in tadpoles was also found to be affected under the influence of the MAT. Changes in the activity of AChE were noticed in all the exposed tadpoles as compared with control Table 2.
TABLE 2: CHANGES IN ACETYLCHOLINESTERASE ACTIVITY OF CONTROL AND EXPOSED TADPOLES OF DUTTAPHRYNUS MELANOSTICTUS FOLLOWING MAT EXPOSURE FOR 5 DAYS, VALUES ARE EXPRESSED AS nmole OF PRODUCT FORMED min/ mg/ PROTEIN
| Parameter/group | C | E1 | E2 | E3 |
| Acetylcholinesterase | 7.81 ± 0.24a | 6.64 ± 0.2b | 5.02 ± 0.25c | 2.15 ± 0.16d |
| Percent change | ----- | -14.98 | -35.72 | -72.471 |
Values are means ± SEM. Means with different superscripts are significantly different (p<0.01), while means with the same superscripts indicate non-significant changes according to one-way ANOVA with Tukey’s post hoc test.
DISCUSSION: Aquatic pollution due to indiscriminate use of pesticides has been acknowledged to be one of an important anthropogenic source against amphibian survival and is known to affect a large fraction of the aquatic ecosystems around the globe. Amphibian species are vulnerable to contaminants because of the greater permeability of the skin and the presence of gills during the larval stage 34, 35. Oxidative metabolism is a regular phenomenon in the cell environment 36, 37, and its controlled limits are governed by biochemical makeup in different aquatic organisms including fish 38, 39, and amphibians 40. The biochemical makeup, constitutes several sections, of which, antioxidant enzymes form an imperative part 36. The antioxidant pool further comprises a cluster of free oxyradical scavenging enzymes, viz. catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx) which are known to be the first line of defense against oxidative stress triggered by arbitrary metabolic process in-vivo 41. Thus, their over and under expressions are known to play a crucial role in the maintenance of the health status of an organism 42.
The present investigation reveals the interference of MAT in oxidative metabolism and thus initiating the cascading changes by the inducement of oxidative stress in Dattaphrynus melanostictus tadpoles. Metabolic breakdown of intoxicated xenobiotics can lead to ROS production at a considerable amount, which is sufficient to compromise the health status of an organism 43. The significant variation in the activity of catalase enzyme witnessed in the present study is in agreement with the studies of David et al., 44 who reported the decline in catalase enzyme activity under the influence of a synthetic pyrethroid, cypermethrin.
In the present study, the elevation of CAT at ‘E1’and depletion at ‘E2’ and ‘E3’ are in agreement with the previously reported studies of Costa et al., 45 who suggested the discrepancies in CAT activity of bullfrog under the influence of Round up®. Variation in catalase activity in the present study did not follow a uniform pattern and was inconsistent with MAT concentration. This may be due to some other extraneous factors which perhaps may not be captured within the boundary of the present study. However, it could be emphasized that the biochemical demolition and repair are the two simultaneous yet individual processes, and this could have accounted for uneven patterns of catalase expression.
Further, the present report on the significant decline in CAT activity in the D. melanostictus tadpoles begs the question of whether the exposed tadpoles could be more tolerant to MAT induced oxidative stress and whether high levels of MAT deactivates CAT or overwhelms the tadpoles to synthesize CAT under mild concentrations. The SOD activity in tadpoles was found to decline under the presence of MAT. Ojha et al., 42 suggested the active role of organophosphorus pesticides in modulating the activity of SOD in a significant fashion. The intracellular environment could be the space for the generation of superoxide anion, which may be the consequence of foreign chemical component participation 46. This generally initiates the active role of SOD upon which it swiftly terminates the harmful radical under the action of body defense mechanism. The constant interaction between elevated levels of superoxide radicals and inadequate magnitude of its counterpart SOD might have resulted in an incessant enzymatic decline in the present investigations.
The glutathione peroxidase activity is known to be a crucial indicator in determining the levels of oxidative stress 47. The elevated activity of GPx at ‘E1’ and its subsequent reduction at ‘E2’ and ‘E3’ may be due to its active role during countering of the free oxyradicals generated during MAT detoxification. The other reason may be perhaps due to the GPx sharing of its substrate H2O2 with CAT. Since the affinity of CAT towards H2O2 is much lesser than GPx 48, and its induction under mild MAT stress could have been a biochemical response of exposed tadpoles. The uniform and irregular trend in decline of SOD and CAT respectively, in the tadpoles exposed to MAT, begs the need for an alternative mechanism to reimburse enzymatic status which was seen as a response under the compensatory act of GPx 49.
This mechanism further convinces the marked increase in GPx activity at E1, during which there was an insignificant change of CAT and significant change in SOD activity. These statements are in support of studies reported by Santos et al., 50 who witnessed the changes in GPx activity in Phyllomedus aiheringii exposed to polluted water. This phenomenon may perhaps be adequate to convince the metabolic synthesis of GPx activity under varying concentrations of MAT.
MDA, being one of the major end products of lipid peroxidation 51, is also thought to be a cause in fabricating the arrest of cell function through elevating the condition of oxidative stress 52. Increased LPO was observed in the present investigation which can be implicated to MAT interaction with cellular metabolism 53, stated that the increased LPO may be due to intervention of a toxic substance such as pesticides, which matches the outline of present condition, wherein, increase in LPO was clearly found to be based on concentration of investigated toxicant MAT. The present findings of increased LPO and decreased antioxidant enzyme activity can reaffirm strong support to suggested hypothesis stating induced LPO could be the consequence of decreased antioxidant enzyme status or vice versa 54.
The acetylcholinesterase activity in tadpoles exposed to different concentrations of MAT is presented in Table 2. From the outcome of the present investigation, it could be noted that, in-vivo exposures to sublethal concentrations of MAT led to a concentration-dependent inhibition of AChE activity. The significant decline in activity of acetylcholinesterase of exposed tadpoles which was observed in the present investigation can be compared with the previously the findings of Lopez-Lopez et al., 55 who reported the significant decline in AChE activity of Girardinichthys viviparous (Bustamante) to untreated domestic wastewater, agricultural runoff and sewage treatment plant effluent. AChE is recognized as a crucial neurotransmitter is known to play a vital act in regulation of cholinergic nervous transmission.
In a report given by Scaps and Borot 56 suggested the inhibition of AChE in the polychaete Nereis diversicolor upon exposure to a carbamate insecticide- carbamyl. Detrimental effects of pesticide mediated inhibition of AChE activity are evident from the current investigation; this is in the background of their sensitivity and urgent need towards their declining population worldwide 57. Reports provided by few authors have suggested digressed locomotory behavior in Neomysis integer 58 and L. vannamei 59 that could be due to inhibition in cholinesterase activity of exposed animals. Several reports have indicated locomotive distress which in turn has been linked to the impaired status of AChE activity in the freshwater oligochaete Lumbriculus variegatus two days after of exposure to azinphos-methyl 60. Therefore, links established in our present study could reaffirm the outcome of AChE inhibition as a biomarker which may further contribute in evaluating the detrimental speculations that could be caused by organophosphorus insecticide MAT on health status and survival ability of D. melanostictus tadpoles. The current study further clearly implicates the possibilities of MAT in causing impaired swimming abilities in tadpoles, following exposure to sublethal concentrations for 5 days.
CONCLUSION: The present investigation suggests that the commercial formulations of MAT can induce oxidative stress through a decline in antioxidant enzyme activity, which may further lead to oxidative damage in D. melanostictus during metamorphosis. MAT was also known to demonstrate antagonistic property against acetylcholinesterase activity in exposed tadpoles. Thus, based on the outcome of the present investigation, it is suggested that necessary precautions should be taken before its use and disposal under the proximity of active amphibian habitats, to conserve the vanishing amphibian species. Further, the assays included in present study validate the feasibility to measure the toxicity of MAT in terms of antioxidant stability, anticholinesterase potentials in the course of regulatory surveillance and monitoring the waters with suspected MAT contamination.
ACKNOWLEDGEMENT: The authors are thankful to the University Grants Commission (UGC), New Delhi, India, for the financial assistance through UGC SAP scheme [No. F.4-18/2015/DSA-I (SAP-II)] and also thankful to Department of Science and Technology for assistance through DST Purse Phase II program.
CONFLICT OF INTEREST: The authors hereby declare no conflict of interest.
REFERENCES:
- Jones DK, Hammond JI and Relyea RA: Very highly toxic effects of endosulfan across nine species of tadpoles: lag effects and family-level sensitivity. Environ ToxicolChem 2009; 28: 1939-45.
- David M and Kartheek RM: Sodium cyanide induced biochemical and histopathological changes in freshwater fish Cyprinuscarpio under sublethal exposure. Int J Toxicol Appl Pharmacol 2014; 4(4): 64-69.
- Kartheek RM and David M: Modulations in hematological aspects of Wistar rats exposed to sublethal doses of fipronil under subchronic duration. Int J Pharm Pharmaceut Sci 2017; 5(3): 177-94.
- Quaranta A, Bellantuono V, Cassano G and Lippe C: Why amphibians are more sensitive than mammals to xenobiotics. PLoS ONE 2009; 4(11): e7699.
- David M, Kartheek RM and Manjunath GP: Acute and sublethal toxicity of chlorpyrifos on developmental stages of Dattaphrynus melanostictus. J Appl Pharm Sci 2018; 8(06): 087-093.
- Touchon JC and Wojdak JM: Plastic hatching timing by red-eyed tree frog embryos interacts with larval predator identity and sublethal predation to affect prey morphology but not performance. PLoS One 2014; 9(6): e100623.
- David M, Shrinivas SJ and Kartheek RM: Biochemical modulations in Duttaphrynus melanostictus tadpoles, following exposure to commercial formulations of cypermethrin: an overlooked impact of extensive cypermethrin use. J Appl Biol Biotechnol 2016, 4(6): 32-037.
- Arribas R, Díaz-Paniagua C and Gomez-Mestre I: Ecological consequences of amphibian larvae and their native and alien predators on the community structure of temporary ponds. Freshwat Biol 2014; 59: 1996-08.
- David M and Kartheek RM: Sodium cyanide induced histopathological changes in kidney of freshwater fish Cyprinus carpio under sublethal exposure. Int J Pharm Chem Biol Sci, 2014; 4(3): 634-39.
- Collins JP and Storfer A: Global amphibian declines: Sorting the hypotheses. Diversit Distribut. 2003; 9: 89-98.
- Knapp RA, Boiano DM and Vredenburg VT: Recovery of a declining amphibian (Mountain Yellow-legged Frog, Rana muscosa) following removal of non-native fish. Biol Conserv 2007; 135: 11-20.
- Hayes T, Haston K, Tsui M, Hoang A, Haeffele C and Vonk A: Atrazine-induced hermaphroditism at 0.1 ppb in American leopard frogs (Ranapipiens): Laboratory and field evidence. Environ Health Persp 2003; 111: 568-75.
- Grayson KL, Bailey LL and Wilbur HM: Life history benefits of residency in a partially migrating pond-breeding amphibian. Ecol 2011: 92: 1236-46.
- Kartheek RM and David M: Assessment of renal toxicity in rats exposed to commercial formulations of fipronil. IntJ Pharm Chem Biol Sci 2017; 7(3): 296-03.
- Brühl CA, Schmidt T, Pieper S and Alscher A: Terrestrial pesticide exposure of amphibians: an underestimated cause for the global decline? Sci Rep 2013; 3: 1135.
- Johnson PTJ, Chase JM, Dosch KL, Hartson RB, Gross JA, Larson DJ, Sutherland DR and Carpenter SR: Aquatic eutrophication promotes pathogenic infection in amphibians. Proc Nat AcadSci USA 2007: 104, 15781-86.
- Groner ML and Relyea RA: A tale of two pesticides: how common insecticides affect aquatic communities. Freshwater Biol 2011; 56: 391-04.
- Boone MD and Semlitsch RD: Interactions of an insecticide with larval density and predation in experimental amphibian communities. Conserv Biol 2001; 15: 228-38.
- Hartman EAH, Belden JB, Smith LM and McMurry ST: Chronic effects of strobilurin fungicides on development, growth and mortality of larval Great Plains toads (Bufo cognatus). Ecotoxicol 2014; 23: 396-03.
- Bridges CM and Semlitsch RD: Variation in pesticide tolerance of tadpoles among and within species of Ranidae and patterns of amphibian decline. Conserv Biol 2000; 14: 1490-99.
- Thiemann GW and Wassersug RJ: Patterns and consequences of behavioral responses to predators and parasites in Rana tadpoles. Bio J Lin So 2000; 71: 513-28.
- Katzenberger M, Hammond J, Duarte H, Tejedo M, Calabuig C and Relyea RA: Swimming with predators and pesticides: how environmental stressors affect the thermal physiology of tadpoles. PLoS ONE 2014; 9: e98265.
- David M and Kartheek RM: Histopathological alterations in the spleen of freshwater fish Cyprinus carpio exposed to sublethal concentration of sodium cyanide. Open Vet J 2015; 5: 1-5.
- Zabin SB, Kartheek RM and David M: Studies on the effect of fipronil on behavioral aspects and protein metabolism of freshwater fish Oreochromis mossambicus. Int J Fish Aquat Stud 2018; 6(3): 221-26.
- David M, Manjunath GP and Kartheek RM: Ameliorative effect of Mentha spicata on dichlorvos-induced oxidative stress in the rat brain. Saudi J Med Pharm Sci 2018; 4(4): 468-76.
- Peltzer PM, Junges CM, Attademo AM., BassóA, Grenón P and Lajmanovich RC: Cholinesterase activities and behavioral changes in Hypsiboas pulchellus (Anura: Hylidae) tadpoles exposed to glufosinate ammonium herbicide. Ecotoxicol 2013; 22: 1165-73.
- David, M and Kartheek, RM: Malathion acute toxicity in tadpoles of Duttaphrynus melanostictus, morphological and behavioural study. The J BasApp Zoo 2015; 72: 1-7.
- Gosner KL: A simplified table for staging anuran embryos and larvae with notes on identification. Herpatol 1960; 16: 183-90.
- Luck HA: Spectrophotometric method for the estimation of catalase In: methods of enzymatic analysis, bergmeyer, H.U. (ed.), Academic Press, New York, 1963; 886-87.
- Kakkar P, Das B and Viswanathan P: A modified method for assay of superoxide dismutase. Ind J Biochem Biophys 1984; 21: 131-32.
- Paglia DE and Valentine WN: Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. Lab Clin Med 1967; 70: 158-69.
- Buege JA and Aust SD: Microsomal lipid peroxidation. In: L Packer (ed). Methods in Enzymology, Acad Press New Jersey 1978; 52: 302-10.
- EllmanGL, Courtney KD, Andres JV and Featherstone RM: A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 1961; 7: 88-95.
- Bridges CM: Tadpole swimming performance and activity affected by acute exposure to sublethal levels of carbaryl. Environ Toxicol Chem 1997; 16: 1935-39.
- Blaustein AR, Romansic JM, Kiesecker JM and Hatch AC: Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers Distr 2003; 9: 123-40.
- Azzam EI, de Toledo SM and Little JB: Oxidative metabolism, gap junctions and the ionizing radiation-induced bystander effect. Nature Oncog 2003; 22: 7050-57.
- Kartheek RM and David M: Fipronil induced modulations in biochemical and histopathological aspects of male Wistar albino rats: A subchronic study. J Environ Biosci 2016; 5(2): 26-32.
- Hagedorn M, McCarthy M, Carter VL and Meyers SA: Oxidative Stress in Zebrafish (Danio rerio) Sperm. PLoS ONE 2012; 7: e39397.
- David M and Kartheek RM: Biochemical changes in liver of freshwater fish Cyprinus carpio exposed to sublethal concentration of sodium cyanide, Indo-Amer J Pharm Res 2014; 4(09): 3669-75.
- Zhang H, Cai C, Shi C, Cao H, Han Z and Jia X: Cadmium-induced oxidative stress and apoptosis in the testes of frog Rana limnocharis. Aquat Toxicol 2012; 122(123): 67-74.
- Kartheek RM and David M: Assessment of fipronil toxicity on Wistar rats: a hepatotoxic perspective. Toxicol Rep 2018; 5: 448-56.
- Ojha A, Yaduvanshi SK, Pant SC, Lomash V and Srivastava N: Evaluation of DNA damage and cytotoxicity induced by three commonly used organophosphate pesticides individually and in the mixture, in rat tissues. Environ Toxicol 2011; 28: 543-52.
- Poljsak B, Šuput D and Milisav I: Achieving the balance between ros and antioxidants: when to use the synthetic antioxidants. Oxidative medicine and cellular longevity. 2013; 956792.
- David M, Munaswamy V, Ramesh H and Marigoudar SR: Impact of sodium cyanide on catalase activity in the exotic freshwater carp, Cyprinus carpio (Linnaeus). Pest Biochem Physiol 2008; 92: 15-18.
- Costa MJ, Monteiro DA, Oliveira-Neto AL, Rantin FT and Kalinin AL: Oxidative stress biomarkers and heart function in bullfrog tadpoles exposed to Roundup original. Ecotoxicol 2008; 17: 153-63.
- Stead D and Park S: Role of Fe superoxide dismutase and catalase in resistance of Campylobacter coli to freeze thaw stress. Appl Environ Microbiol 2000; 66: 3110-12.
- Blokhina O, Virolainen E and Fagerstedt KV: Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann Bot Lond.) 2003; 91: 179-94.
- David M and Kartheek RM: In-vivo studies on hepato-renal impairments in freshwater fish Cyprinus carpio following exposure to sublethal concentrations of sodium cyanide. Environ Sci Pollut Res Int 2016; 23: 722-33.
- Wang D, Li F, Chi Y and Xiang J: Potential relationship among three antioxidant enzymes in eliminating hydrogen peroxide in penaeid shrimp. Cel Stre Ch 2012; 17: 423-33.
- Santos TG, Melo R, Costa e Silva D, Nunes M and Franco JFJ: Assessment of water pollution in the Brazilian Pampa BIOME using stress biomarkers in tadpoles of the leaf frog Phyllomedus aiheringii (Anura: Hylidae) Peer J PrePrints 2015; 3: e776v1.
- Hodgson E: A textbook of modern toxicology, John Wiley & Sons, Hoboken, NJ, USA 2004.
- Lushchak VI: Environmentally induced oxidative stress in aquatic animals. Aquat Toxicol 2011; 101: 13-30.
- Sharma DK and Ansari BA: Effects of deltamethrin on CAT, LPO and GSH in tissues of zebrafish, Daniorerio. Res J Environ Toxico 2013; 7: 38-46.
- Grim JM, Hyndman KA, Kriska T, Girotti AW and Crockett EL: Relationship between oxidizable fatty acid content and level of antioxidant glutathione peroxidases in marine fish. The J Exper Biol 2011; 214: 3751-59.
- Lopez-Lopez E, Sedeno-Diaz JE and Perozzi F: Lipid peroxidation and acetylcholinesterase activity as biomarkers in the Black Sailfin Goodeid, Girardinichthys viviparous (Bustamante) exposed to water from Lake Xochimilco (Mexico). Aquat Ecosys Health Manag 2006; 9: 379-85.
- Scaps P and Borot O: Acetylcholinesterase activity of the polychaete Nereis diversicolor: effects of temperature and salinity. Comp Biochem Physiol Part C: Pharmacol Toxicol Endocrinol 2000; 125: 377-83.
- Peltzer PM, Lajmanovich RC, Attademo AM, Junges CM, Cabagna-Zenklusen MC, Repetti MR, Sigrist ME and Beldoménico H: Effect of exposure to contaminated pond sediments on survival, development, and enzyme and blood biomarkers in veined. Ecotoxicol Environ Saf 2013; dx.doi.org/10.1016/j.ecoenv.2013.09.010i.
- Roast SD, Widdows J and Jones MB: Disruption of swimming in the hyper benthic mysid Neomysis integer (Peracarida: Mysidacea) by the organophosphate pesticide chlorpyrifos. Aquat Toxicol 2000; 47: 227-41.
- García-de la Parra LM, Bautista-Covarrubias JC, Riverade la Rosa N, Betancourt-Lozano M and Guilhermino L: Effects of methamidophos on acetylcholinesterase activity, behavior, and feeding rate of the white shrimp (Litopenaeus vannamei). Ecotoxicol Environ Saf 2006; 65: 372-80.
- Kristoff G, Guerrero NR and Cochón AC: Inhibition of cholinesterase activity by azinphosmethyl in two freshwater invertebrates: Biomphalaria glabrata and Lumbriculus variegates. Toxicol 2006; 222: 185-94.
How to cite this article:
Kartheek RM, David M, Manjunath GP, Lokeshkumar P and Mahantesh D: Assessment of malathion induced toxicity in Dattaphrynus melanostictus tadpoles: a biochemical investigation. Int J Pharmacognosy 2019; 6(6): 216-23. doi link: http://dx.doi.org/10.13040/IJPSR. 0975-8232.IJP.6(6).216-23.
This Journal licensed under a Creative Commons Attribution-Non-commercial-Share Alike 3.0 Unported License.
Article Information
4
216-223
621
602
English
IJP
R. M. Kartheek, M. David *, G. P. Manjunath, P. Lokeshkumar and D. Mahantesh
Department of Zoology, JSS Banashankari Arts Commerce and SK Gubbi Science College, Dharwad, Karnataka, India.
mdavid.kud@gmail.com
30 May 2019
23 June 2019
25 June 2019
10.13040/IJPSR.0975-8232.IJP.6(6).216-23
30 June 2019